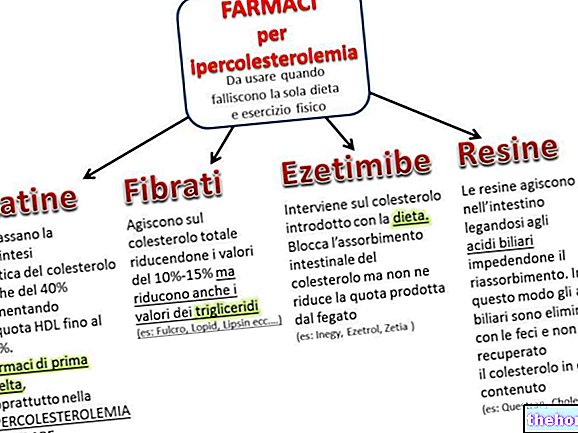
Elusolendite tsükliline järjepidevus leiab paljunemisnähtustes seoseid järgnevate põlvkondade vahel.
Paljundamine toimub evolutsioonilise skaala erinevatel tasanditel, erinevates taime- ja loomariikide tagajärgedes, erinevates elusliikides, kasutades nii erinevaid mehhanisme, mis üksi õigustavad tervet traktaati.
Paljunemisnähtuste esimene klassifikatsioon peab eristama üherakulisi ja mitmerakulisi organisme, kuna ainult esimeses langeb rakkude jagunemine kokku paljunemisega.
Mitmerakulises võib paljunemine olla agamiline või seksuaalne (või gamiline).
Agamiline paljunemine, suhteliselt harvem, põhineb mitoosi mehhanismil, nii et liigi varieeruvus on pigem usaldatud mutatsioonide korduvale esinemisele.
Samuti on olemas erinevad mehhanismid, näiteks strobiliseerimine, eostumine jne, samas kui taimedest leiame põllumajanduses hästi tuntud taastavaid vorme (pistikud, kihilisus jne).
Kõrgemate vormide kõige levinum paljunemismehhanism on aga seksuaalne, mis vastab meioosi ilmnemisele, sugurakkude moodustumisele ja nende liitumisele sügootis (viljastumine).
Primitiivsetel liikidel ei ole sugurakud morfoloogiliselt diferentseerunud: sel juhul räägime isogametiast. Siiski on alati kaks sugurakkude seeriat, mis on tähistatud sümbolitega (+) ja (-), ja viljastumine saab toimuda ainult "vastandmärgiga sugurakkude kohtumisest": seetõttu on bioloogiline erinevus, mis pole veel morfoloogiliselt väljendatud.
Evolutsioonilise skaala edenedes ilmneb morfoloogiline ja funktsionaalne diferentseerumine, mille puhul emaste sugurakkude tüüp on tavaliselt varustatud rikkaliku varumaterjaliga (deutoplasma või vasikas, mis tagab embrüo arengu kuni see on metaboolselt sõltumatu) ja teatud tüüpi isased sugurakud, kellel on liikumisvõime naissoost. Gametes on alati haploidsed ja tulenevad meioosist. Nende sulandumisel tekib diploidne sügoot.
Meioosi ja sügootide vahel võib kulgeda haploidsete rakkude põlvkondade seeria, nii nagu sügootide ja meiooside vahel võib kuluda rida diploidsete rakkude põlvkondi, kusjuures generatsioonitsüklite erinevaid sorte väljendab põlvkondade vaheldumine.
Diplonte liiki (millel on diploidne organism) iseloomustab gameetiline meioos: meioos toodab otseselt sugurakke, mis ühendades koheselt taastavad diploidse oleku. See on metaaslaste, sealhulgas inimese, levinud juhtum.
Gametogenees
Võttes arvesse paljunemist gametilise meioosiga, nagu see esineb inimesel, proovime selgitada, kuidas meioos sobib gametogeneesi (sugurakkude moodustumine).
Embrüonaalses arengus, meeste ja naiste gametogeneesis (mida nimetatakse spermatogeneesiks ja oogeneesiks), eristatakse varakult keha moodustavaid rakke (somaatiline joon) ja sugurakke tootvaid rakke (iduliin). Suguliini esialgseid rakke nimetatakse protogoonideks. Näärmete diferentseerumisel isas- või naissoost mõttes toimub sugurakkude diferentseerumine vastavalt spermatogooniaks ja ovogooniaks.
Vaadates spermatogeneesi, näeme, et spermatogoonias on rida rakupõlvesid, mis jätkuvad kogu elu. Ainult osa pidevalt toodetud spermatogooniast erineb tavalisest mitootilisest tsüklist ja selle asemel algab meiootiline tsükkel.
Sugurakk, milles meioos algab (reduplikatsioon ja seejärel esimene jagunemine), nimetatakse esimese järgu spermatotsüütideks; selle jagunemisel tekib kaks teise järgu spermatotsüüti, mis teise jagunemisega tekitavad kokku neli spermatiiti.
Me võime lõpetada kromosoomikomplekti vähendamise esimese astme spermatotsüütide 4n -st (pärast reduplikatsiooni on neli kromatiili iga homoloogipaari kohta) teise järgu spermatotsüütide 2n -ni ja spermatiidide n -ni, nagu juba uurides näha. meioos, mida seega järeldame. Seepärast on spermatiidid juba haploidsed, kuid nad ei ole veel küpsed sugurakud. Haploidset tüüpi raku struktuurist muudab funktsionaalne küpsemine (nimetatakse spermiohistogeneesiks) spermatiidid spermatosoidideks, see tähendab küpseteks isasrakkudeks.
Naiste gametogeneesis (või oogeneesis) on mitmeid erinevusi. Esiteks on ettevalmistatavate sugurakkude arv palju väiksem. Hinnanguliselt valmistatakse inimliigi emase sugunäärmetes umbes 5 x 105 ovogooni; neist ainult umbes 400 on huvitatud folliikulite küpsemisest ja sellele järgnevast masendusest, tsüklis, mis tavaliselt mõjutab ainult ühte folliikulit kuus umbes 35 -aastase viljaka perioodi jooksul.
Mõlemast soost valmistatud sugurakkude erinev arv vastab juba mainitud funktsioonide ja käitumise erinevusele: spermatosoidid on väikesed, liikuvad ja arvukad seoses muna otsimise vajadusega ja selle leidmise väikese tõenäosusega; munad on suured, inertsed ja vähesed, seoses funktsiooniga tagada embrüole varumaterjal ja neile sisemise viljastamise abil pakutav kaitse (loomulikult peab ka munarakke olema rohkem).
Vajadus varustada sugurakke varumaterjaliga vastab "oogeneesis" meioosi peatamise faasi olemasolule, mille käigus kromosoomid on osaliselt despiraliseeritud. Seejärel jälgime niinimetatud "sulgede kromosoome". Ekstraversioonid tuvastavad tunnused, milles deutoplasma sünteesi eest vastutavad geenid on alla surutud.
Emaste sugurakkude jaoks vajalik väiksem arv vastab ka asjaolule, et neljast meioosi tekitatud haploidsest rakust saab ainult üks kogu reservmaterjali ja saab sugurakk, ülejäänud kolm (polotsüüdid või polaarkehad), mis sisaldavad ainult kromosomaalne materjal, ei saa tekitada sügoote ja embrüoid ning on määratud taanduma.
Väetamine
Viljastamist, see tähendab isaste ja emaste sugurakkude kohtumist, saab läbi viia väga erineval viisil.Loomariigis jälgime üleminekut väliselt viljastamisest (sugurakud, mis on avatud mis tahes keskkonnaohule ja seega tingimata väga arvukad mõlemast soost) sisemisele viljastamisele, millega vanemlik hoolitsus on veelgi seotud imetajate ainevahetussuhtega ema ja loote vahel .
Kui vastassugupoole sugurakkude kohtumine on toimunud, peab väetamine toimuma, tagades kaks tingimust: spetsiifilisuse ja ainulaadsuse. See tähendab, et tuleb tagada, et spermatosoid on samast liigist nagu muna ja et kui esimene siseneb, ei sisene teised.
Spetsiifilisuse tagavad akrosoomi ja munaraku pinna biokeemilised omadused. Tegelikult räägitakse "viljastamisvahendite" ja "viljastumisvastaste ainete" vahelistest reaktsioonidest, mille spetsiifilisus on võrreldav ensüümide ja substraadi kokkupuutega.
Väetamise ainulaadsuse tagab munaraku pinna struktuuri muutmine ("kortikaalne reaktsioon"), mis algab kohe esimese spetsiifilise väetise / antifertilisiini reaktsiooni hetkel; pärast seda reaktsiooni muudetakse munaraku membraani, nii et teised sinna jõudnud spermatosoidid ei suuda enam spetsiifilist viljastamisreaktsiooni käivitada.
Pärast viljastamist jääb spermatosoidi saba munarakust väljapoole, samas kui kromosoomimaterjal tungib sellesse. See, mida nimetatakse "isaseks pronukleuseks", ühineb munaraku "emase pronukleusega", moodustades seega tsügooti diploidse tuuma.