Nukleiinhapped on suure bioloogilise tähtsusega keemilised ühendid; kõik elusorganismid sisaldavad nukleiinhappeid DNA ja RNA kujul (vastavalt desoksüribonukleiinhape ja ribonukleiinhape). Nukleiinhapped on väga olulised molekulid, kuna neil on esmane kontroll kõigi organismide põhiliste eluprotsesside üle.
Kõik viitab sellele, et nukleiinhapped on mänginud identset rolli alates primitiivse elu esimestest vormidest, mis suutsid ellu jääda (näiteks bakterid).
Elusorganismide rakkudes leidub DNA peamiselt kromosoomides (jagunevates rakkudes) ja kromatiinis (intertsneetilistes rakkudes).
Seda leidub ka väljaspool tuuma (eriti mitokondrites ja plastiidides, kus see täidab oma funktsiooni teabekeskusena osa või kogu organelli sünteesiks).
RNA seevastu esineb nii tuumas kui ka tsütoplasmas: tuumas on see kontsentreeritum tuumas, tsütoplasmas aga polüsoomides.
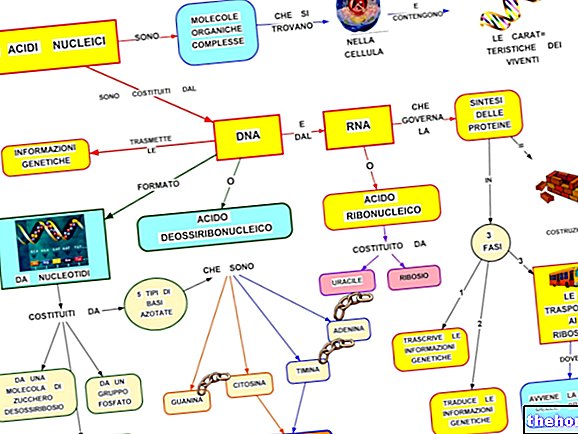
Nukleiinhapete keemiline struktuur on üsna keeruline; neid moodustavad nukleotiidid, millest igaüks (nagu oleme näinud) koosneb kolmest komponendist: süsinikhüdraat (pentoos), lämmastikalus (puriin või pürimidiin) ja fosforhape.
Nukleiinhapped on seega pikad polünukleotiidid, mis tulenevad nukleotiidideks nimetatud üksuste ühendamisest. Erinevus DNA ja RNA vahel seisneb pentoosis ja aluses. Pentoosi on kahte tüüpi, üks igale nukleiinhappe tüübile:
1) Riboos RNA -s;
2) Dessosiriboos DNA -s.
Aluste osas peame eristust kordama; pürimidiini aluste hulka kuuluvad:
1) tsütosiin;
2) tümiin, esineb ainult DNA -s;
3) Uracil, esineb ainult RNA -s.
Puriinalused seevastu koosnevad:
1) Adeniin
2) Guaniin.
Kokkuvõttes leiame DNA -st: tsütosiin - adeniin - guaniin - tümiin (C -A -G -T); samas kui RNA -s on meil: tsütosiin - adeniin - guaniin - uratsiil (C -A -G -U).
Kõigil nukleiinhapetel on polünukleotiidne lineaarne ahela struktuur; teabe eripära annab aluste erinev järjestus.
DNA struktuur
DNA ahela nukleotiidid on seotud ester -sidemega fosforhappe ja pentoosi vahel; hape on seotud nukleotiidpentoosi süsinikuga 3 ja järgmise süsinikuga 5; nendes sidemetes kasutab ta kahte oma kolmest happerühmast; ülejäänud happerühm annab molekulile happelise iseloomu ja võimaldab moodustada sidemeid aluseliste valkudega .
DNA -l on kahekordne spiraalstruktuur: kaks üksteist täiendavat ahelat, millest üks "läheb alla" ja "teine" läheb üles. "Sellele paigutusele vastab" paralleelsete "ahelate mõiste, see tähendab paralleelsed, kuid vastupidiste suundadega. ühel küljel algab üks ahelatest sidemega fosforhappe ja pentoosi süsiniku 5 vahel ning lõpeb vaba süsinikuga 3; samal ajal kui täiendava ahela suund on vastupidine. Samuti näeme, et nende kahe ahela vahel tekivad vesiniksidemed ainult puriinaluse ja pürimidiini vahel ja vastupidi, st adeniini ja tümiini vahel ning tsütosiini ja guaniini vahel ja vastupidi; AT -paaris on kaks vesiniksidet, GC -paaris aga kolm sidet. See tähendab, et teisel paaril on suurem stabiilsus.
DNA reduplikatsioon
Nagu juba mainitud seoses intertsüneetilise tuumaga, võib DNA olla "autosünteesi" ja "allosünteesi" faasis, see tähendab vastavalt osaleda enda paaride (autosüntees) või "muu aine" (RNA: allosüntees) sünteesil. See on jagatud kolme faasi, mida nimetatakse G1, S, G2. Faasis G1 (kus G võib olla algne kasv) sünteesib rakk tuuma DNA kontrolli all kõik, mis on vajalik selle ainevahetuseks. Faasis S (kus S tähistab sünteesi, st uue tuuma DNA sünteesi) toimub DNA reduplikatsioon. G2 faasis jätkab rakk kasvu, valmistudes järgmiseks jagunemiseks.
VAATAME LÜHIDALT S ETAPIS SISALDUNUD NÄHTUSI
Esiteks võime kujutada kahte paralleelset ahelat nii, nagu oleksid need juba "despiraliseeritud". Alustades ühest äärmusest, purunevad sidemed aluspaaride (A - T ja G - C) vahel ja kaks üksteist täiendavat ahelat liiguvad teineteisest eemale (sobib "välgu" avanemise võrdlus). Siinkohal ensüüm ( DNA-polümeraas) "voolab" mööda igat üksikut ahelat, soodustades sidemete tekkimist seda moodustavate nukleotiidide ja karüoplasmas levinud uute (varem "ATP vabastatud energiaga" aktiveeritud) nukleotiidide vahel. Uus timína on tingimata seotud iga adeniiniga ja nii edasi, moodustades järk -järgult igast ahelast uue topeltketi.
DNA-polümeraas näib mõjuvat in vivo ükskõikselt kahele ahelale, olenemata "suunast" (3 kuni 5 või vastupidi). Sel viisil, kui kogu algne kahekordne DNA ahel on kaetud, on kaks topeltahelat täpselt sama mis originaal. Mõiste, mis seda nähtust määratleb, on "poolkonservatiivne reduplikatsioon", kus "reduplikatsioon" koondab kvantitatiivse kahekordistamise ja täpse koopia tähendused, samas kui "poolkonservatiivne" tuletab meelde asjaolu, et iga uue kahekordse DNA ahela puhul üks ahel on neointeetiline.
DNA sisaldab geneetilist teavet, mille ta edastab RNA -le; viimane edastab selle omakorda valkudele, reguleerides seega raku ainevahetusfunktsioone.Järelikult on kogu ainevahetus otseselt või kaudselt tuuma kontrolli all.
DNA -st leitud geneetilise pärandi eesmärk on anda rakule spetsiifilisi valke.
Kui võtame need paarikaupa, annab neli alust 16 võimalikku kombinatsiooni, see tähendab 16 tähte, mis ei ole kõigi aminohapete jaoks piisav. Kui võtame need hoopis kolmikutena, on 64 kombinatsiooni, mida võib tunduda liiga palju, kuid mis tegelikult on kõik kasutusel, kuna teadus on avastanud, et erinevaid aminohappeid kodeerib rohkem kui üks kolmik. Seetõttu on meil tõlge nukleotiidide lämmastikaluste neljast tähest 21 aminohappele; enne tõlkimist on aga c "transkriptsioon", mis jääb endiselt "neljatäheliseks", see tähendab geneetilise teabe liikumine DNA neljast tähest RNA nelja täheni. arvestades, et häbeliku (DNA) asemel on c "uratsiil (RNA).
Transkriptsiooniprotsess toimub siis, kui ribonukleotiidide, ensüümide (RNA-polümeraas) ja ATP molekulides sisalduva energia juuresolekul avaneb DNA ahel ja sünteesitakse RNA, mis on selles osas sisalduv geneetiline teave. avatud ahel.
RNA -d on kolm peamist tüüpi ja need kõik pärinevad tuuma -DNA -st:
- RNAm (sõnumitooja)
- RNAr (ribosomaalne)
- RNAt või RNA -d (ülekantavad või lahustuvad)