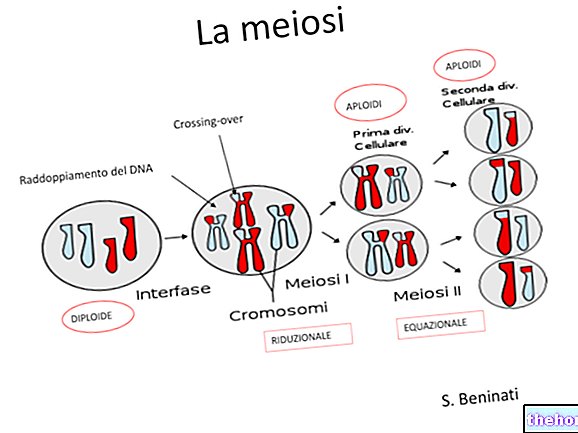
ÜHENDUS (VÕI "KOKKUVÕTMINE" VÕI "LINKAGE")
Siiani oleme käsitlenud Mendeli dihübriidi (või polühübriidi) ristumist, eeldades, et erinevaid alleelipaare leidub tegelikult erinevatel homoloogilistel kromosoomipaaridel. Kuid kromosomaalsete paaride arv, ehkki liigiti erinev, varieerub kitsastes piirides (vähesed liigid ulatuda umbes saja kromosoomini), samas kui geenide arvu võib lugeda kümnetesse tuhandetesse.
See, et Mendeli oma katseteks valitud tegelased eraldusid kõik üksteisest sõltumatult (segadusse ajamata seega arvutusi fenotüüpide jaotuse kohta polühübriidristi F2 -s), oli õnnelik juhus. Kui külgnevatest lookustest leiti kaks paari alleele, kehtib seadus järgiks seda nimetataks ühinguseaduseks.
Teades, et väga paljudel tegelastel on oma lookus ühes kromosoomis ja et need on kromosomaalsed paarid, mis eralduvad meioosis iseseisvalt, on hästi mõistetav, kui sageli juhtub, et kaks paari tähemärki, kui nad on seotud vanema kromosoomiga samuti sugurakkudes ja seega organismis, kuhu ta toob oma geneetilise materjali.
Seega näeme, et "assotsiatsioon" kujutab endast erandit, mis pole kaugeltki harv, Mendeli kolmandas seaduses sätestatud iseseisvuse suhtes.
VAHETUS VÕI "ÜLEMINE" JA KOMBINATSIOON
Meioosist rääkides oleme märkinud, et geneetilise materjali segunemisel on kaks erinevat momenti: üks on sugurakkude kromosoomide segregatsioon ja seda täheldas Mendel.
Teine hetk, mis tegelikult eelneb, on see, kus iga paari homoloogiliste kromosoomide neli kromatiidi vahetavad vastastikku identseid tunnuseid. Pärast seda vahetust on kaks samas kromosoomis seostatud tegurit sugurakkudes sõltumatud. Tõenäosus et vahetus toimub, on esimese lähendamise korral proportsionaalne kromosoomi pikkusega ja pikemates kromosoomides võib olla isegi rohkem kui vahetus.
Seda nähtust saab tuvastada tsütoloogiliselt, jälgides mikroskoobi all piisavat arvu meioose.
Uuesti sobivuse määr on kiirus, millega kaks vanemliku põlvkonnaga seotud märki F2-s erinevalt rekombineeruvad.
Kui need kaks lookust on absoluutselt külgnevad, on tõenäosus, et kiasm eraldab need, praktiliselt null. Rekombinatsiooni määr on: n ° rekombinandid. Kui kaks lookust on kahel erineval kromosoomil, on rekombinatsiooni määr 0,5 (võrdne tõenäosus kahe P -põlvkonnaga ühendatud märgi puhul sattuda juhuslikult F2 -sse). Seega võib rekombinatsiooni kiirus varieeruda vahemikus 0,0 kuni 0,5. Kromosoomi väikeste vahemaade korral on vahemaa ja rekombinatsiooni kiirus otseselt proportsionaalsed. Pikemate vahemaade puhul on võimalik, et kahe lookuse vahel toimub kaks vahetust. Nüüd on selge, et kaks tegurit, mida eraldab kaks vahetust, on taas seotud. siinkohal on selge, et proportsionaalsus lookuste kauguse ja rekombinatsiooni tõenäosuse vahel on kadunud.
Samas kromosoomis seostatud lookused moodustavad „assotsiatsioonirühmad”. Väga kaugetel lookustel võib olla selline vahetusega eraldumise tõenäosus, et nad käituvad sõltumatuna, kuid igaüks neist seotakse madalama rekombinatsioonikiirusega vahepealsete lookustega.
Kui assotsiatsioonirühma paljude geenipaaride vahel on rekombinatsiooni määrad teada, võib alata geneetiliste kaartide koostamine. Pidades meeles, et kahe geeni (a ja b) vahelist kaugust väljendab rekombinatsiooni määr ja et a kaugus kolmandast geenist c võib olla kas summa või erinevus selle kaugusest b -st, on võimalik rekonstrueerida vastastikuste vahemaade kaart, mis on selle assotsiatsioonigrupi, st selle kromosoomi geneetiline kaart.
Peame nüüd üldiselt kaaluma mõningaid mõisteid, mis piiravad genotüübiliste tegelaste fenotüüpilist avaldumist.
Kõigepealt räägime läbitungivuse ja väljendusvõime mõistetest ning seejärel pöörame erilist tähelepanu geenitoimingu reguleerimise nähtustele.
PETETRANCE
Geeni läbitungivus näitab selle võimet avalduda fenotüübis. Läbitungimist mõõdetakse statistiliselt, lugedes fenotüüpide esinemissageduse, mis näitavad seda iseloomu 100 genotüübist, mis seda sisaldavad. Tunnus 0,7 läbitungivusega on tunnus, mis esineb fenotüüpiliselt 70% selle genotüübilisest sagedusest.
VÄLJENDAVUS
Ekspressiivsus on fenotüüpse manifestatsiooni astme kvantitatiivne hindamine.
Geenitegevuse reguleerimine
Rakud toodavad kõiki oma ensüüme ja valke sama kiirusega ja samal ajal.Näiteks Escherichia coli rakke saab varustada energia- ja süsinikuaatomitega laktoosdisahhariidist, kuna nad suudavad need lagundada glükoosiks ja galaktoosiks tänu beeta-galaktosidaasi ensüümile. Normaalse E. coli korral, mis võib sisaldada laktoosi, seal on ligikaudu 3000 beeta-galaktosidaasi molekuli, mis võrdub 3% selle raku valkudega; laktoosi puudumisel on bakteriraku kohta ainult üks beeta-galaktosidaasi molekul. Galaktosidaasi sünteesitakse uutest mRNA molekulidest, kui seda saab kasutada. Ensüümirikkad E. coli mutantsed tüved on teada isegi siis, kui laktoosi pole: need mutandid on normaalsete rakkudega võrreldes ebasoodsas olukorras, kuna nad on sunnitud tarbetult kulutama energiat ja materjale, et toota ensüümi, mis jääb ilma substraadita. Aineid, mis põhjustavad ensüümi hulga suurenemist, nagu laktoosi puhul, nimetatakse indutseerijateks, ensüüme aga indutseeritavateks. Teised ained kutsuvad esile ka teatud viisil teatud ensüümide tootmise. Ka näiteks E. coli puhul, mis on võimeline ehitama kõiki oma süsinikku ja ammooniumi (NH3) sisaldavaid aminohappeid, blokeerib teatud aminohappe (näiteks histidiin) olemasolu söötmes kõigi ensüümide tootmist, mis on seotud aminohappe enda biosüntees: nende ensüümide kohta öeldakse, et need on allasurutavad. Bakterirakkudes lammutatakse mRNA molekulid varsti pärast nende moodustumist ja seetõttu tähendab mRNA tootmise kontrollimine samal ajal ensümaatilise sünteesi juhtimist aega.
OPERON
Et selgitada, kuidas bakterirakk on võimeline kontrollima oma ensüümide tootmist, sõnastasid Jacob ja Monod operoni hüpoteesi; operoni moodustavad mitmed geenid, mis on funktsionaalselt seotud ja katkematult joondatud piki DNA lõiku.Operon koosneb kolmest erinevat tüüpi geenist: promootor, kust algab mRNA moodustumine; operaator, kus teostatakse kontrolli; üks või mitu struktuurgeeni, mis kodeerivad ensüüme või muid valke. Beeta-galaktosidaasi süsteemis sisaldab operon lisaks beeta-galaktosidaasile ka kahte muud geeni, mis struktuurselt kodeerivad muud laktoosi metabolismis osalevad ensüümid. Need geenid asuvad üksteise kõrval ja transkribeeritakse üksteise järel mööda sama DNA heeliksit üheks mRNA molekuliks. Selliselt toodetud mRNA molekulid on aktiivsed väga lühikest aega, misjärel need hävivad spetsiifiliste ensüümide poolt.
Operoni aktiivsust kontrollib omakorda teine geen - regulaator, mis võib samuti olla operonist kaugel: see regulaator kodeerib valku, mida nimetatakse repressoriks ja mis näib seonduvat operaatorgeeni DNA -ga. promootori ja struktuurigeenide vahel tegelikult blokeerib mRNA tootmist.
Repressorit juhitakse omakorda ja juhtimine toimub "signaal" aine abil. Indutseeritavate ensüümide puhul on see aine indutseerija. Indutseerija seondub repressormolekuliga, muutes selle kuju nii, et see ei suuda enam DNA -ga kohaneda: antud juhul kuna promootori ja struktuurigeenide vahel pole repressorit , võib repressor moodustada mRNA molekule ja neist valgu molekule. Kui indutseerijavarud on ammendunud, saab regulaator uuesti kontrolli, mis peatab uue mRNA, seega uute valkude tootmise. Beeta-galaktosidaasi süsteemis on indutseerijaks laktoos või sellele väga sarnane aine. Derivaat: need liitub repressoriga, inaktiveerides selle, et võimaldada ensüümide biosünteesi. Represseeritavate ensüümide puhul toimib "signaalina" toimiv aine tuumakompressorina: repressor on aktiivne ainult siis, kui see on kombineeritud tuumakompressoriga. Histidiinisüsteemis, mis hõlmab tosinat erinevat ensüümi, on see aminohape koos selle tRNA -ga, corepressor histidiin.
ALLOSTERILISED SUHTED
Allosteerilised koostoimed, mis hõlmavad ensüümi inaktiveerimist selle kuju muutmise teel, pakuvad raku metaboolse aktiivsuse reguleerimiseks erinevat viisi. Allosteerilised koostoimed võimaldavad täpsemat kontrolli kui operaatori induktiiv -repressorsüsteem, kuid ei saavuta kasulikku tulemust, kui välistada antud aine biosüntees esimesest etapist - mRNA tootmisest.
KONTROLLI SÜSTEEMID EUCARIUSES
On mõningaid fakte, mis panevad arvama, et taimedel ja loomadel on operatsioonisüsteemiga sarnane ja opereeriv süsteem. Nende organismide kromosoomid erinevad sügavalt E. coli ja teiste prokarüootide kromosoomidest. Mitoosi mehhanism on selline, et antud taime või looma iga rakk omab kogu teavet
viljastatud munarakkudes esinev geneetika. Seetõttu jääb enamik mis tahes spetsialiseeritud raku geene kogu raku eluea jooksul ebaefektiivseks. Nende rakkude DNA on alati seotud valkudega. Seega on võimalik, et geenide allasurumine eukarüootides nõuab just seda seost DNA ja valkude vahel.





















-cos-cause-e-come-superarla.jpg)
