Toimetanud dr Giovanni Chetta
Alates biokeemiast kuni biomehaanikani

Rakuväline maatriks (MEC)
ECM (rakuväline maatriks) kirjeldus, ehkki vähe sellest, mida me täna teame, on oluline selgroo ja kehahoia muutuste tähtsuse paremaks mõistmiseks tervises.
Iga rakk, nagu iga mitmerakuline elusorganism, peab oma elutähtsate funktsioonide täitmiseks ja ellujäämiseks "tundma" ja suhtlema oma keskkonnaga. Mitmerakulises organismis peavad rakud koordineerima erinevat käitumist nagu inimeste kogukonnas.

Üldiselt kirjeldatakse ECM -i, mis koosneb mitmest suurest biomolekulide klassist:
- Struktuursed valgud (kollageen ja elastiin)
- Spetsiaalsed valgud (fibrilliin, fibronektiin, laminiin jne)
- Proteoglükaanid (agrekaanid, sündekaanid) ja glusaminoglükaanid (hüaluroonid, kondroitiinsulfaadid, heparaansulfaadid jne)
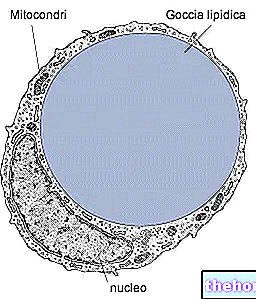
Struktuursete valkude hulgas, s.t kollageen moodustavad loomariigis kõige enam esindatud glükoproteiinide perekonna. Need on ECM -is kõige rohkem esinevad valgud (kuid mitte kõige olulisemad) ja on õige sidekoe (kõhre, luu, fastsia, kõõlused, sidemed) põhikomponendid.
Kollageene sünteesivad enamasti fibroblastid, kuid epiteelirakud suudavad neid ka sünteesida.
Kollageenikiud suhtlevad pidevalt tohutu hulga teiste ECM -i molekulidega, mis moodustavad raku eluks olulise bioloogilise järjepidevuse. Seotud kollageenid fibrillides omavad domineerivat rolli struktuuride moodustamisel ja säilitamisel, mis on võimelised vastu pidama peaaegu pingejõududele. elastset kollageeni toodetakse ja metaboliseeritakse uuesti mehaanilise koormuse mõjul ning selle viskoelastsetel omadustel on suur mõju inimese kehahoiakule.
Kollageenkiududel on tänu oma PG / GAG kattele (proteoglükaanid / glükoosaminoglükaanid) biosensor- ja biojuhtivusomadused. Me teame tegelikult, et igasugune mehaaniline jõud, mis on võimeline tekitama struktuurseid deformatsioone, rõhutab molekulidevahelisi sidemeid, tekitades kerge elektrivoo, st piesoelektrilise voolu (Athenstaedt, 1969). Seetõttu omab kolmemõõtmeline ja kõikjal esinev kollageenivõrk ka bioelektriliste signaalide juhtimist ruumi kolmes dimensioonis, mis põhineb kollageenkiudude ja rakkude suhtelisel paigutusel aferentses suunas (ECM-st rakkudeni) või vastupidi. vastupidi efferentne.
Kõik see kujutab endast reaalajas MEC-raku sidesüsteemi ja sellised elektromagnetilised biosignaalid võivad viia oluliste biokeemiliste muutusteni, näiteks juhul, kui „luu osteoklastid ei suuda piesoelektriliselt laetud luu seedida“ (Oschman, 2000).
Lõpetuseks tuleb rõhutada, et rakk toodab üllatuslikult pidevalt ja märkimisväärsete energiakuludega (umbes 70%) materjali, mis tuleb tingimata väljutada ainsa protokollageeni (kollageeni bioloogilise eelkäija) säilitamise teel konkreetsetes vesiikulites ( Albergati, 2004).
Valdav osa selgroogsete kudedest eeldab kahe olulise omaduse - tugevuse ja elastsuse - samaaegset olemasolu. Tõeline võrgustik elastsed kiud, mis asub nende kudede ECM -i sees, võimaldab pärast tugevaid tõmbeid naasta esialgsetesse tingimustesse. Elastsed kiud on võimelised suurendama elundi või selle osa venivust vähemalt viis korda. Elastsete kiudude vahele on paigutatud pikad, elastsed kollageenikiud, mille täpne ülesanne on piirata "liigset deformatsiooni kudede tõmbamise teel." Elastiin on elastsete kiudude põhikomponent ja seda leidub eriti rikkalikult elastsete omadustega veresoontes ( moodustab üle 50% aordi kogumassist), sidemetes, kopsudes ja nahas. Silelihasrakud ja fibroblastid on selle lähteaine tropoelastiini peamised tootjad.
ECM sisaldab suurt hulka (ja veel täpselt määratlemata) spetsialiseeritud mitte-kollageenvalke, millel on tavaliselt spetsiifilised sidumissaidid teiste ECM molekulide ja rakupinna retseptorite jaoks. Sel moel toimib nende valkude iga komponent nii sarnaste kui ka erinevate molekulide vaheliste kontaktide "võimendajana", luues lõpmatu biokeemilise võrgustiku, mis on võimeline genereerima, moduleerima, muutma ja paljundama isegi vahemaa tagant miljoneid ja miljoneid biokeemilist teavet (ja energiat).
Rakuvälise maatriksi oluline "spetsialiseeritud valk" on fibronektiin, suure molekulmassiga glükoproteiin, mida leidub kõigil selgroogsetel.Fibronektiin näib olevat võimeline mõjutama rakkude kasvu, rakkudevahelist adhesiooni ja koos ECM -iga, rakkude migratsiooni erinevatel viisidel (rakk võib liikuda kuni 5 cm päevas - Albergati, 2004) jne. Kõige tuntum III tüüpi isovorm seondub integriinidega . Viimased on transmembraansete valkude perekond, mis toimivad mehhanoretseptoritena: nad muudavad valikuliselt ja moduleeritaval viisil mehaanilisi tõmbeid ja tõukeid rakusisest ECM -ist ja vastupidi, kutsudes esile tsütoplasmas mitmeid reaktsioone, mis hõlmavad tsütoskeleti ja muud valgud, mida nad reguleerivad rakkude adhesiooni, kasvu ja migratsiooni (Hynes, 2002).
Glükoosaminoglükaanid (GAGS) ja proteoglükaanid (PG) moodustavad sidekudedes määratletud kõrgelt hüdreeritud geelitaolise aine, milles paiknevad ja imbuvad fibrillaarsed valgud. See polüsahhariidgeeli vorm võimaldab ühelt poolt ECM -il vastu pidada märkimisväärsetele survejõududele ja teiselt poolt võimaldab toitainete, metaboliitide ja hormoonide kiiret, pidevat ja selektiivset difusiooni vere ja kudede vahel.
Glükoosaminoglükaanide polüsahhariidahelad on mahult liiga jäigad, et neid polüpeptiidahelatele tüüpiliste kompaktsete kerakujuliste struktuuride sisse voltida, lisaks on need väga hüdrofiilsed. Nendel põhjustel (ja ilmselt ka muudel meile tundmatutel) kipuvad GAG -d eeldama ülimalt head konformatsiooni. hõivavad oma massi suhtes suure mahu ja moodustavad seega isegi väikestes kontsentratsioonides märkimisväärse koguse geeli. Suur hulk negatiivseid laenguid (GAG -d esindavad kõige rohkem anioonseid rakke, tavaliselt sulfaaditud, loomarakkude poolt toodetud) meelitab ligi palju katioone; nende hulgas on ülekaalus Na +, mis annab kogu osmootse võimsuse ja hoiab ECM -is tohutu koguse vett. Sel viisil tekivad tursed (turgorid), mis võimaldavad ECM -il astuda vastu isegi olulistele survejõududele (tänu sellele võib näiteks puusa kõhre füsioloogilistes tingimustes suurepäraselt vastu pidada mitmesaja atmosfääri rõhule).
Sidekoes moodustavad GAG-d vähem kui 10–12% kogukaalust, kuid tänu oma omadustele täidavad need paljusid rakuväliseid ruume, moodustades erineva suuruse ja tihedusega elektrilaengutega hüdreeritud geeli poorid, toimides seega selektiivselt võtmepunktid või "serverid", mille kaudu reguleeritakse MEC -i molekulide ja rakkude liiklust nende suuruse, kaalu ja elektrilaengu alusel.
Hüaluroonhape (hüaluronaan, hüaluronaat) on GAG-idest ehk lihtsaim. Eksperimentaalsed ja molekulaarbioloogilised andmed kinnitavad, et sellel on luude ja liigeste tasemel oluline roll seoses märkimisväärse survega. ECM-i ruumide täitmine embrüonaalse arengu ajal : see loob rakkude vahele tühjad ruumid, kuhu nad hilisemates etappides rändavad (Albergati, 2004).
ECM ei erita kõiki PG -sid, mõned on plasmamembraanide lahutamatud komponendid (Alberts, 2002).
Seetõttu võib rakuvälist maatriksit pidada väga keerukaks võrgustikuks, milles valgud, PGS ja GAG-d täidavad lugematuid funktsioone, sealhulgas struktuurset tuge ja iga koe ja orgaanilise tegevuse reguleerimist. Globaalset rakulist homöostaasi tuleks käsitleda mehhanismide kompleksina, mis võivad pärineda ja areneda rakus või väljaspool ECM -i; viimasel juhul võib rakk esindada vahe- või lõppsihtmärki. Rakuvälised komponendid on lisaks rakuliste karkasside füüsilistele tugistruktuuridele ka tegelikud saidid nii endotsellulaarse keskkonna kui ka elundite ja süsteemide elutähtsate protsesside käivitamiseks, arendamiseks ja lõpetamiseks. Oleme silmitsi lõpmatu biokeemilise võrgustikuga, mis on võimeline genereerima, moduleerima, varieerima ja levitama isegi eemalt miljoneid ja miljoneid teavet.
Iga keharakk suhtleb pidevalt ECM -iga nii mehaanilises kui ka keemilises ja energeetilises aspektis, avaldades "dramaatilist" mõju kudede staatilisele ja dünaamilisele arhitektuurile. P. A. Bacci sõnul kujutab interstitsiaalne maatriks tõeliselt elutähtsate reaktsioonide ema, kohta, kus toimuvad ennekõike aine ja energia vahetused. Kõik koed on omavahel ühendatud ja funktsionaalselt integreeritud mitte suletud, vaid avatud süsteemides; nende vahel toimub pidev vahetus, mis võib toimuda nii lokaalselt kui ka süsteemselt, kasutades ära biokeemilisi, biofüüsikalisi ja elektromagnetilisi sõnumeid, st kasutades erinevaid energiavorme.
Nagu F. G. Albergati kinnitab, kujutavad rakk ja rakuväline maatriks kahte näiliselt eraldiseisvat maailma, mis peavad tingimata kogu eluaja vältel toimima korrektselt ja sünergiliselt. See nõuab erakordset signaalide jada, millele järgneb sama uskumatu molekulaarbioloogiliste tegevuste seeria.
Muud artiklid teemal "Rakuväline maatriks - struktuur ja funktsioonid"
- Skolioosi ravi
- Skolioos - põhjused ja tagajärjed
- Skolioosi diagnoosimine
- Skolioosi prognoos
- Sidekude ja sidekoe
- Ühendusriba - funktsioonid ja funktsioonid
- Rüht ja pinge
- Inimese liikumine ja tuuletõmbuse tähtsus
- Õigete põlve- ja hambumustugede tähtsus
- Idiopaatiline skolioos - müüdid hajutamiseks
- Skolioosi ja raviprotokolli kliiniline juhtum
- Ravi tulemused Kliinilise juhtumi skolioos
- Skolioos kui loomulik hoiak - Bibliograafia











.jpg)